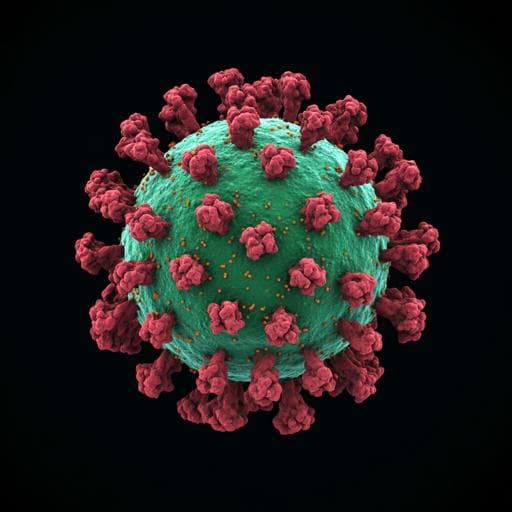
Structural insights into the modulation of coronavirus spike tilting and infectivity by hinge glycans
D. Chmielewski, E. A. Wilson, et al.
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.
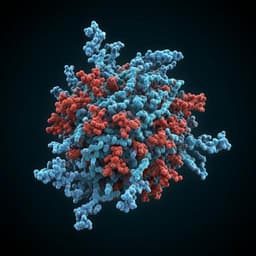
Targeting the coronavirus SARS-CoV-2: computational insights into the mechanism of action of the protease inhibitors lopinavir, ritonavir and nelfinavir
G. Bolcato, M. Bissaro, et al.
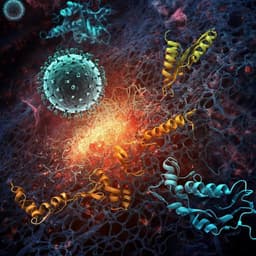
Histones released by NETosis enhance the infectivity of SARS-CoV-2 by bridging the spike protein subunit 2 and sialic acid on host cells
W. Hong, J. Yang, et al.

Three-Dimensional Structural Insights Have Revealed the Distinct Binding Interactions of Agonists, Partial Agonists, and Antagonists with the µ Opioid Receptor
Z. Li, J. Liu, et al.
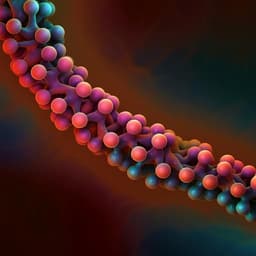
Conversion of levoglucosan into glucose by the coordination of four enzymes through oxidation, elimination, hydration, and reduction
Y. Kuritani, K. Sato, et al.


