BiologyNature Communications
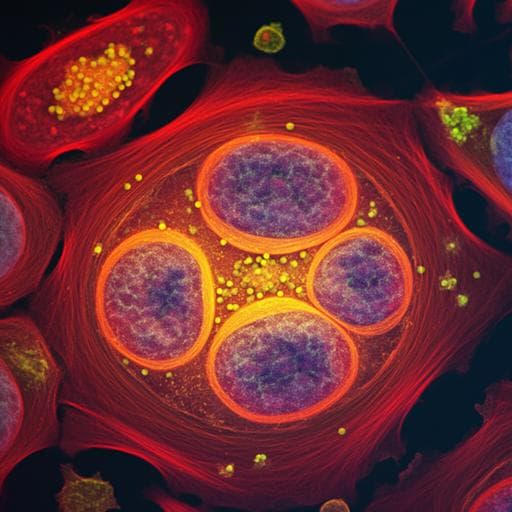
High-dimensional super-resolution imaging reveals heterogeneity and dynamics of subcellular lipid membranes
K. Zhanghao, W. Liu, et al.
Discover the groundbreaking Spectrum and Polarization Optical Tomography (SPOT) technique developed by researchers including Karl Zhanghao and Wenhui Liu, which allows for real-time monitoring of subcellular lipid dynamics in live cells. This innovative method provides insights into lipid heterogeneities during cell division across multiple organelles.
Related Publications
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.

Engineering and Technology
Fast non-line-of-sight imaging with high-resolution and wide field of view using synthetic wavelength holography
F. Willomitzer, P. V. Rangarajan, et al.
Engineering and Technology
Bottom-up construction of low-dimensional perovskite thick films for high-performance X-ray detection and imaging
S. Dong, Z. Fan, et al.

Biology
In vivo volumetric imaging of calcium and glutamate activity at synapses with high spatiotemporal resolution
W. Chen, R. G. Natan, et al.
Engineering and Technology
Overtone photothermal microscopy for high-resolution and high-sensitivity vibrational imaging
L. Wang, H. Lin, et al.


