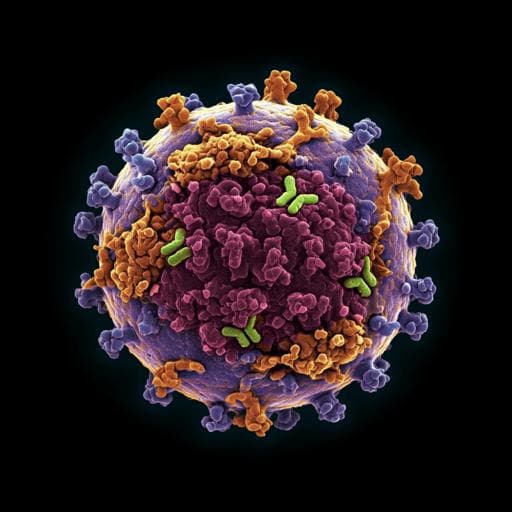
Cryo EM structures map a post-vaccination polyclonal antibody response to canine parvovirus
S. R. Hartmann, A. J. Charnesky, et al.
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.
Links between fecal microbiota and the response to vaccination against influenza A virus in pigs
M. Borey, F. Blanc, et al.

The cryo-EM structure of the bd oxidase from *M. tuberculosis* reveals a unique structural framework and enables rational drug design to combat TB
S. Safarian, H. K. Opel-reading, et al.

A novel modelling approach to quantify the response of dairy goats to a high-concentrate diet
M. Taghipoor, M. Delattre, et al.

Efficacy of early PET-CT directed switch to carboplatin and paclitaxel based definitive chemoradiotherapy in patients with oesophageal cancer who have a poor early response to induction cisplatin and capecitabine in the UK: a multi-centre randomised controlled phase II trial
S. Mukherjee, C. N. Hurt, et al.


