
Targeting senescence induced by age or chemotherapy with a polyphenol-rich natural extract improves longevity and healthspan in mice
S. Zumerle, M. Sarill, et al.
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.

Psychological and ethical issues raised by genomic in paediatric care pathway, a qualitative analysis with parents and childhood cancer patients
M. Droin-mollard, S. D. Montgolfier, et al.

AV futures or futures with AVs? Bridging sociotechnical imaginaries and a multi-level perspective of autonomous vehicle visualisations in praxis
R. Martin
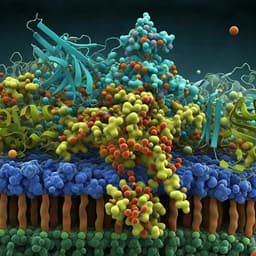
MFRP is a molecular hub that organizes the apical membrane of RPE cells by engaging in interactions with specific proteins and lipids
A. Tworak, R. Smidak, et al.

Leveraging Large Language Models for Precision Monitoring of Chemotherapy-Induced Toxicities: A Pilot Study with Expert Comparisons and Future Directions
O. R. Sarrias, M. P. M. D. Prado, et al.


