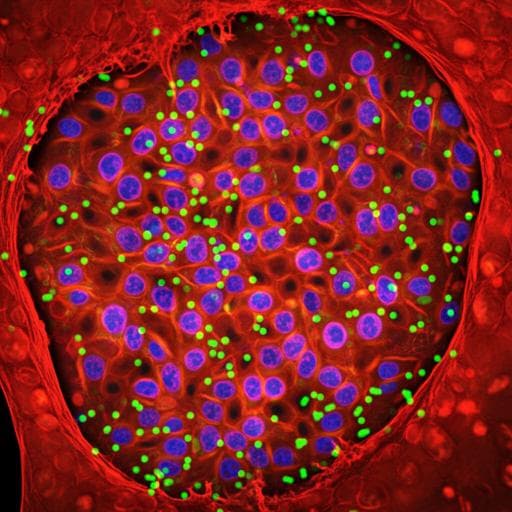
Medicine and HealthNature Methods
Image-seq: spatially resolved single-cell sequencing guided by in situ and in vivo imaging
C. Haase, K. Gustafsson, et al.
Explore the pioneering advancements in spatial and temporal understanding of tissue function through Image-seq technology, developed by leading researchers including Christa Haase and Karin Gustafsson. This innovative approach merges single-cell analysis with spatial organization, unveiling crucial insights in leukemia biology and beyond.
Related Publications
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.
Biology
Single-cell RNA sequencing reveals shared and distinct immune responses in Kawasaki disease and COVID-19
X. Liu, T. Luo, et al.
Medicine and Health
speedingCARs: accelerating the engineering of CAR T cells by signaling domain shuffling and single-cell sequencing
R. Castellanos-rueda, R. B. D. Roberto, et al.
Veterinary Science
Single-cell RNA sequencing reveals the cellular and molecular heterogeneity of treatment-naïve primary osteosarcoma in dogs
D. T. Ammons, L. S. Hopkins, et al.
Medicine and Health
High resolution mapping of the tumor microenvironment using integrated single-cell, spatial and in situ analysis
A. Janesick, R. Shelansky, et al.


