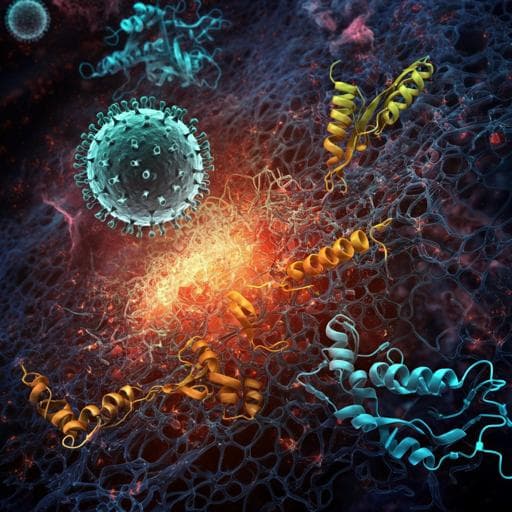
Histones released by NETosis enhance the infectivity of SARS-CoV-2 by bridging the spike protein subunit 2 and sialic acid on host cells
W. Hong, J. Yang, et al.
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.

Evolution of the SARS-CoV-2 spike protein in the human host
A. G. Wrobel, D. J. Benton, et al.

The COVID-AGICT study: COVID-19 and advanced gastro-intestinal cancer surgical treatment. A multicentric Italian study on the SARS-CoV-2 pandemic impact on gastro-intestinal cancers surgical treatment during the 2020. Analysis of perioperative and short-term oncological outcomes
G. Giuliani, F. Guerra, et al.
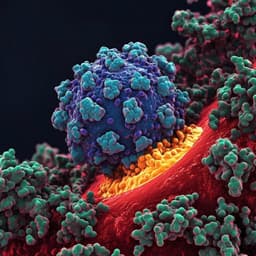
Broad host range of SARS-CoV-2 and the molecular basis for SARS-CoV-2 binding to cat ACE2
L. Wu, Q. Chen, et al.
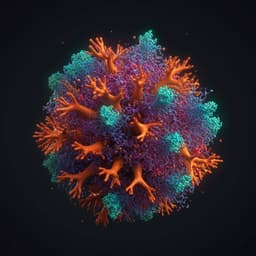
Analysis of the N-glycosylation profiles of the spike proteins from the Alpha, Beta, Gamma, and Delta variants of SARS-CoV-2
D. Wang, J. Baudys, et al.


