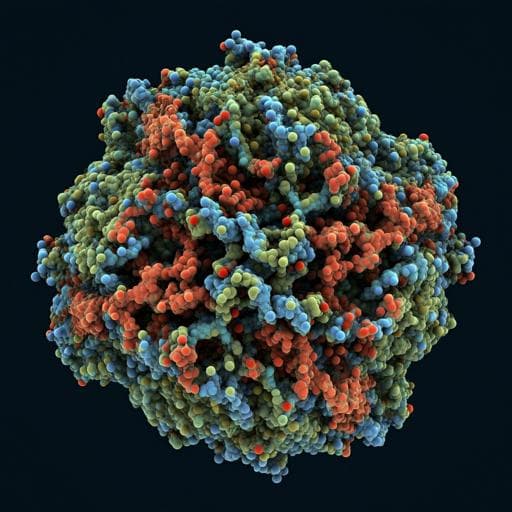
Atomic structure of a nudivirus occlusion body protein determined from a 70-year-old crystal sample
J. R. Keown, A. D. Crawshaw, et al.
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.

Auditory Discrimination-A Missing Piece of Speech and Language Development: A Study on 6-9-Year-Old Children with Auditory Processing Disorder
A. Guzek and K. Iwanicka-pronicka

Reliability of body composition assessment using A-mode ultrasound in a heterogeneous sample
M. Miclos-balica, P. Muntean, et al.
A synergistic impact of body mass index and gamma gap on heart failure and mortality rate among older patients with coronary artery disease: a prospective study with 10-year follow-up
X. Liu, Y. Zheng, et al.
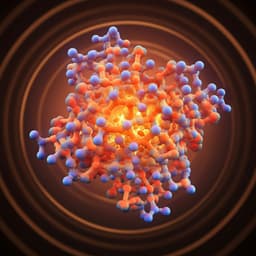
A Computational Biology Study on the Structure and Dynamics Determinants of Thermal Stability of the Chitosanase from Aspergillus fumigatus
Q. Wang, S. Liu, et al.


