
A deeper understanding of system interactions can explain contradictory field results on pesticide impact on honey bees
D. Breda, D. Frizzera, et al.
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.
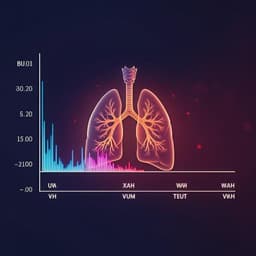
Impact of the severity of restrictive spirometric pattern on nutrition, physical activity, and quality of life: results from a nationally representative database
S. J. Chung, H. I. Kim, et al.
Impact of baseline adipose tissue characteristics on change in adipose tissue volume during a low calorie diet in people with obesity—results from the LION study
D. Junker, M. Wu, et al.

A pilot study: the impact of clinic-provided transportation on missed clinic visits and system costs among teenage mother-child dyads
L. Allan-blitz, A. Samad, et al.

The evolution of social-ecological system interactions and their impact on the urban thermal environment
B. Chen, F. Kong, et al.


