Medicine and Healthnpj Vaccines
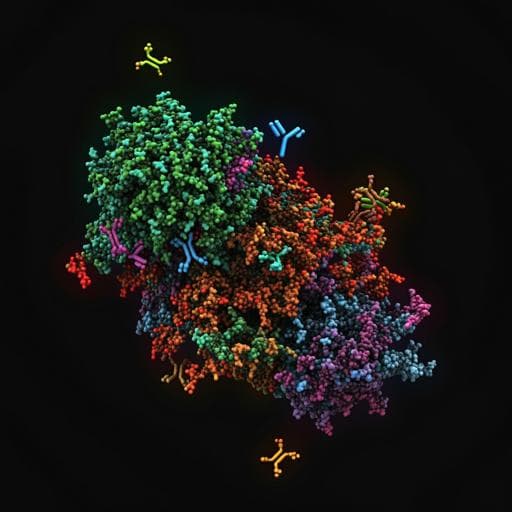
Potent neutralization of SARS-CoV-2 variants by RBD nanoparticle and prefusion-stabilized spike immunogens
M. C. Miranda, E. Kepl, et al.
This groundbreaking study explores how RBD-NP immunogens, refined with mutations from SARS-CoV-2 variants like B.1.351 and P.1, can adapt to Omicron strains. The authors reveal that these innovative vaccines can elicit strong neutralizing antibody responses against variants, demonstrating rapid design and stability, making them promising candidates for a broadly protective coronavirus vaccine platform.
Related Publications
Explore these studies to deepen your understanding
Adjacent work that informs or extends this paper's methodology and findings.
Medicine and Health
Broad ultra-potent neutralization of SARS-CoV-2 variants by monoclonal antibodies specific to the tip of RBD
H. Ma, Y. Guo, et al.
Medicine and Health
Immunogenicity and protection of a variant nanoparticle vaccine that confers broad neutralization against SARS-CoV-2 variants
J. Logue, R. M. Johnson, et al.
Medicine and Health
Potent neutralizing nanobodies resist convergent circulating variants of SARS-CoV-2 by targeting diverse and conserved epitopes
D. Sun, Z. Sang, et al.
Medicine and Health
Histones released by NETosis enhance the infectivity of SARS-CoV-2 by bridging the spike protein subunit 2 and sialic acid on host cells
W. Hong, J. Yang, et al.


